Митохондрии (МТ) – одно из самых интересных мне направлений исследований. Объединение митохондрий с другой клеткой в ходе эндосимбиоза около 1,6 млрд лет назад стало основной всех многоклеточных эукариотов со сложной структурой. Предположительно митохондрии произошли от клеток, напоминающих α-протеобактерии.
Лучшее обзорное исследование последнего времени по митохондриях – работа Вернера Кулбрандта «Структура и функция митохондриальных белковых комплексов мембраны ». Если вы знаете английский язык и интересуетесь устройством этих органелл, то настоятельно рекомендую к прочтению. Эта статья так хороша, что может быть смело главой хорошего учебника по молекулярной биологии. Сначала я хотел перевести всю статью, но это бы заняло непростительно много времени и оторвало бы от других дел. Поэтому ограничусь тезисами и картинками. Периодически разбавляя все своими мыслями.
Митохондрия кодирует сама только 13 белков, не смотря на наличие отдельной от клетки ДНК (мтДНК) и всего «производственного» цикла по транскрипции белков. Изолированная митохондрия какое-то время может сохранять композицию и функционировать.
Внешняя мембрана пористая и позволяет веществам из цитоплазмы проходить через нее. Внутренняя мембрана плотная, для ее пересечения нужны транспортные белки [Гилберт Линг обоснованно не согласен], непрерывность барьера позволяет иметь внутренней мембране электрохимический потенциал в -180 mV. У матрикса довольно большой pH (7,9-8). Еще раз углублюсь в Линга. Щелочной (выше 7) pH способствует более развернутой конформации белков . Высокий pH нарушает водородные и солевые связи, делая поляризованные CO и NH доступными молекулам воды, там самым усиливая дипольный момент всей внутриклеточной воды и связывая ее. В этом ключе наличие мембраны нужно не для «удержания» протоплазмы внутри клетки (это делают сами белки при высоком pH), а для наличия потенциала.
мтДНК находится в нуклеотидах, которых примерно 1000 на клетку. Белковая плотность матрикса довольна высокая (до 500 мг/мл), что близко к кристаллизованным белкам.
Внутренняя мембрана образует инвагинации, называемые кристами, которые глубоко проникают в матрикс. Кристы определяют третий «отсек» митохондрий – просвет крист (cristae lumen). Мембраны кристы содержат большинство, если не все, полностью «собранные» комплексы цепи переноса электронов и АТФ-синтазы. Просвет кристы содержит большое количество маленького растворимого белкового переносчика электронов (цитохром с). Митохондриальные кристы, таким образом, основное место биологической конверсии энергии во всех не фотосинтетических эукариотах.
С кристами тоже много всего интересного. Оптические свойства кристы влияют на распространение и генерацию света в тканях. Я даже встречал идеи о том, что поверхность крист подобна (предположение) поверхностям топологических изоляторов (подразумевалась суперпроводимость без диссипации заряда).
Теперь немного внимания на комплекс II. Вы помните, что жир (кето) делает упор в метаболизме на FADH2 и комплекс II. Они восстанавливают пару CoQ, в какой-то момент окисленного CoQ не хватает для транспортировки электронов на комплекс III и образует обратный поток электронов на комплекс I с образованием супероксида. При долгом HFLC-питании комплекс I будет обратимо разрушен, при этом это нормальная физиологическая оптимизация.
Еще прошу вас заметить, что комплекс II не выкачивает протоны. Что у нас рассеивает протоновый градиент, нарушает фосфорилирование и стимулирует сжигание жиров на тепло? Правильно, стресс холода. Термогенез связан с метаболизмом через комплекс, который не выкачивает протоны, тем самым не давая дополнительных протонов для АТФ-синтазы. Можно только удивляться как замечательно у нас продуман организм.
Места крепления крист (cristae junctions) – маленькие круглы отверстия примерно 25 нм диаметром. В митохондриях всех организмов есть система MICOS (mitochondria contact site and cristae to outer membrane), сборка из пяти мембранных и одного растворимого белков, прикрепляющих кристы к наружной мембране.
В клетках с повышенной потребностью в энергии, такие как скелетные и сердечные мышцы, кристы плотно заполняют большую часть объема митохондрии. В тканях с меньшими потребностями в энергии, таких как печень и почки, кристы находятся не так плотно по отношению друг к другу. Остается больше места в матрице для биосинтетических ферментов.
Митохондриальная F1-F0 АТФ синтаза является самым заметным белковым комплексом кристы. АТФ синтаза – это древняя наномашина, которая использует электрохимический протновых градиент вокруг внутренней мембраны для создания АФТ посредством вращательного катализа
. Протоны, двигающиеся через F0 комплекс мембраны, вращают ротор из 8 (у млекопитающих) или 10 (у дрожжей) с-узлов. Центральный стебель передает крутящий момент c-ротора каталитической головке F1, где АТФ образуется из АДФ и фосфата через последовательность конформационных изменения. Периферийный стебель предотвращает непродуктивное вращение Головы F1 против комплекса F0.
Многие годы считалось, что АТФ синтаза случайным образом располагается на внутренней мембране. Но оказалось, что АТФ синтаза располагается двойными рядами
. Причем линейные ряды АТФ синтазы – фундаментальный атрибут всех живых митохондрий.
Ряды АТФ синтазы располагаются в основном вдоль хребтов крист. Димеры изгибают липидный бислой и как следствие само-организуются в ряды. Когда у митохондрий дрожжей выбивали узлы e и g АФТ синтазы, то штамм рос на 60% медленней диких собратьев, и потенциал мембран их митохондрий был снижен вдвое. У АФТ синтазы прокариотов недостает нескольких узлов, связанных с димерами, ряды димеров не были найдены у бактерий и архей. Кристы и ряды димеров АФТ синтазы, таким образом, являются адаптацией к большим энергетическим потребностям организма.
Протоновый градиент вокруг внутренней мембраны создается тремя крупными мембранными комплексами, известными как комплекс I, комплекс III и комплекс IV (см. рисунок 2). Комплекс I кормится электронами из NADH, высвобождаемая при передаче электрона энергия выкачивает четыре протона. Комплекс III получает электрон от восстановленного хинола и передает его носителю электронов (цитохрому с), выкачивая в процессе один протон. Комплекс IV получает электрон из цитохрома с и передает его молекулярному кислороду, выкачивая 4 протона за каждую молекулу кислорода, превращенную в воду. Комплекс II не выкачивает протоны, напрямую передавая электроны хинолу. Как перенос электронов из NADH в хинол связан с транслокацией протонов пока не ясно. Комплекс I – крупнее III и IV вместе взятых.
Основным белком просвета кристы является цитохром с, который переносит электрон из комплекса III в комплекс IV. Если цитохром с высвобождается в цитоплазму клетки, то вызывает апоптоз
.
Старение – фундаментальный и плохо понимаемый процесс всех эукариотов. Исследовали старение митохондрий на грибах Podospora anserina
, которые живут всего 18 дней. В нормальной митохондрии кристы проникают глубоко в матрикс. Для этого нужны ряды димеров АТФ синтазы и MICOS комплекс у мест крепления крист. С возрастом кристы начинают все ближе подходить к поверхности мембраны, димеры АФТ синтазы превщаются в мономеры, и все заканчивается высвобождением цитохрома с и клеточной смертью.
Транспорт электронов создает супероксид в комплексах I и III. Это побочный продукт метаболизма. Одновременно необходимый и смертельно опасный. Во время старения деление (fission) начинает превалировать над сращением (fussion). Это не дает поврежденным митохондриям «спастись» путем сращения и ускоряет неизбежное.
Митохондрия.
Митохондрия
- состоящая из двух мембран органелла толщиной около 0,5 мкм.
Энергетическая станция клетки; основная функция - окисление органических соединений и использование, освобождающейся при их распаде энергии в синтезе молекул атф (универсальный источник энергии для всех биохимических процессов).
По своему строению они представляют собой цилиндрические органеллы, встречающиеся в эукариотической клетке в количестве от нескольких сот до 1-2 тысяч и занимающие 10-20 % её внутреннего объёма. Сильно варьируют так же размеры (от 1 до 70 мкм) и форма митохондрий. При этом ширина этих частей клетки относительно постоянна (0,5-1 мкм). Способны изменять форму. в зависимости от того, в каких участках клетки в каждый конкретный момент происходит повышенное потребление энергии, митохондрии способны перемещаться по цитоплазме в зоны наибольшего энергопотребления, используя для движения структуры клеточного каркаса эукариотической клетки.
Красавица митохондрия в 3д представлении)
Альтернативой множеству разрозненных небольших митохондрий, функционирующих независимо друг от друга и снабжающих атф небольшие участки цитоплазмы, является существование длинных и разветвлённых митохондрий, каждая из которых может энергетически обеспечивать отдалённые друг от друга участки клетки. вариантом такой протяжённой системы может также являться упорядоченное пространственное объединение множества митохондрий (хондриом или митохондрион), обеспечивающее их кооперативную работу.
Особенно сложно этот тип хондриома устроен в мышцах, где группы гигантских разветвлённых митохондрий связаны друг с другом с помощью межмитохондриальных контактов (ммк). Последние образованы плотно прилегающими друг к другу наружными митохондриальными мембранами, в результате чего межмембранное пространство в этой зоне имеет повышенную электронную плотность (много отрицательно заряженных частиц). Особенно обильно ммк представлены в клетках сердечных мышц, где они связывают множественные отдельные митохондрии в согласованную работающую кооперативную систему.
Структура.
Наружная мембрана.
Наружная мембрана митохондрии имеет толщину около 7 нм, не образует впячиваний и складок, и замкнута сама на себя. на наружную мембрану приходится около 7 % от площади поверхности всех мембран клеточных органелл. Основная функция - отграничение митохондрии от цитоплазмы. Наружная мембрана митохондрии состоит из двойного жирового слоя (как и у клеточной мембраны) и пронизывающих его белков. Белки и жиры в равных пропорциях по массе.
Особую роль играет порин
-
каналообразующий белок.
Он формирует в наружной мембране отверстия диаметром 2-3 нм, через которые могут проникать небольшие молекулы и ионы. Крупные молекулы могут пересекать наружную мембрану только посредством активного транспорта через транспортные белки митохондриальных мембран. Наружная мембрана митохондрии может взаимодействовать с мембраной эндоплазматического ретикулума; это играет важную роль в транспортировке липидов и ионов кальция.
Внутренняя мембрана.
Внутренняя мембрана образует многочисленные гребневидные складки -
кристы,
существенно увеличивающие площадь ее поверхности и, например, в клетках печени составляет около трети всех клеточных мембран. характерной чертой состава внутренней мембраны митохондрий является присутствие в ней кардиолопина
-
особый сложный жир, содержащего сразу четыре жирные кислоты и делающего мембрану абсолютно непроницаемой для протонов (положительно заряженных частиц).
Ещё одна особенность внутренней мембраны митохондрий - очень высокое содержание белков (до 70 % по весу), представленных транспортными белками, ферментами дыхательной цепи, а также крупными ферментами комплексами производящими атф. Внутренняя мембрана митохондрии в отличие от внешней не имеет специальных отверстий для транспорта мелких молекул и ионов; на ней, на стороне, обращенной к матриксу, располагаются особые молекулы ферменты производящие атф, состоящие из головки, ножки и основания. При прохождении через них протонов происходит создание атф.
В основании частиц, заполняя собой всю толщу мембраны, располагаются компоненты дыхательной цепи. наружная и внутренняя мембраны в некоторых местах соприкасаются, там находится специальный белок-рецептор, способствующий транспорту митохондриальных белков, закодированных в ядре, в матрикс митохондрии.
Матрикс.
Матрикс
- ограниченное внутренней мембраной пространство. В матриксе (розовом веществе) митохондрии находятся ферментные системы окисления пирувата жирных кислот, а так же ферменты типа трикарбоновых кислот (цикл дыхания клетки). Кроме того, здесь же находится митохондриальная днк, рнк и собственный белоксинтезирующий аппарат митохондрии.
пируваты (соли пировиноградной кислоты)
- важные химические соединения в биохимии. Они является конечным продуктом обмена веществ глюкозы в процессе ее расщепления.
Митохондриальная днк.
Несколько отличий от днк ядерной:
- митохондриальная днк – кольцевая, в отличии от ядерной днк, которая упакована в хромосомы.
- между различными эволюционными вариантами митохондриальной днк одного вида невозможен обмен сходными участками.
И поэтому вся молекула изменяется только путем медленного мутирования в течение тысячелетий.
- мутации кода в митохондриальных днк могут возникать независимо от ядерной днк.
Мутация ядерного кода днк возникает в основном при делении клетки, но митохондрии делятся независимо от клетки, и могут получать мутацию кода отдельно от ядерной днк.
- сама структура митохондриальной днк упрощена, т.к. многие составные процессы чтения днк утеряны.
- транспортные рнк имеют одинаковое строение. но рнк- митохондрий учавствуют только в синтезе митохондриальных белков.
Имея собственный генетический аппарат, митохондрия обладает и собственной белоксинтезирующей системой, особенностью которой в клетках животных и грибов являются очень маленькие рибосомы.
Функции.
Энергообразование.
Основной функцией митохондрий является синтез атф - универсальной формы химической энергии в любой живой клетке.
Данная молекула может образовываться двумя путями:
- путем реакции, в которых энергия освобождающаяся на определенных окислительных этапах брожения запасается в виде атф.
- благодаря энергии, выделяющейся при окислении органических веществв в процессе клеточного дыхания.
Митохондрии реализуют оба эти пути, первый из которых характерен для начальных процессов окисления и происходит в матриксе, а второй завершает процессы энергообразования и связан с кристами митохондрий.
При этом своеобразие митохондрий как энергообразующих органелл эукариотической клетки определяет именно второй путь генерации атф, получивший название «хемиосмотического сопряжения».
В целом весь процесс энергообразования в митохондриях может быть разбит на четыре основные стадии, первые две из которых протекают в матриксе, а две последние - на кристах митохондрий:
1) Превращение поступивших из цитоплазмы в митохондрию пируват (конечный продукт расщепления глюкозы) и жирных кислот в ацетил-коа;
ацетил-коа
– важное соединение в обмене веществ, используемое во многих биохимических реакциях. его главная функция – доставлять атомы углерода (с) с ацетил-группой (ch3 co) в цикл клеточного дыхания, чтобы те были окислены с выделением энергии.
клеточное дыхание
-
совокупность биохимических реакций, протекающих в клетках живых организмов, в ходе которых происходит окисление углеводов, жиров и аминокислот до углекислого газа и воды.
2) Окисление ацетил-соа в цикле клеточного дыхания, ведущее к образованию надн;
НАДН
–
кофермент, выполняет функцию переносчика электронов и водорода, которые принимает от окисляемых веществ.
3) Перенос электронов с надн на кислород по дыхательной цепи;
4) Образование атф в результате деятельности мембранного атф-создающего комплекса.
АТФ- синтетаза.
АТФ-синтетаза
–
станция по производству молекул АТФ.
В структурно-функциональном плане АТФ-синтетаза состоит из двух крупных фрагментов, обозначаемых символами F1 и F0. Первый из них (фактор сопряжения F1) обращён в сторону матрикса митохондрии и заметно выступает из мембраны в виде сферического образования высотой 8 нм и шириной 10 нм. Он состоит из девяти субъединиц, представленных пятью типами белков. Полипептидные цепи трёх субъединиц α и стольких же субъединиц β уложены в похожие по строению белковые глобулы, которые вместе образуют гексамер (αβ)3, имеющий вид слегка приплюснутого шара.
Субъединица
– это структурный и функциональный компонент какой либо частицы
Полипептиды
- органические соединения, содержащие от 6 до 80-90 аминокислотных остатков.
Глобула
– состояние макромолекул, в котором колебание звеньев мало.
Гексамер
– соединение содержащее 6 субъедениц.
Подобно плотно уложенным долькам апельсина, последовательно расположенные субъединицы α и β образуют структуру, характеризующуюся симметричность относительно угла поворота 120°. В центре этого гексамера находится субъединица γ, которая образована двумя протяжёнными полипептидными цепями и напоминает слегка деформированный изогнутый стержень длиной около 9 нм. При этом нижняя часть субъединицы γ выступает из шара на 3 нм в сторону мембранного комплекса F0. Также внутри гексамера находится минорная субъединица ε, связанная с γ. Последняя (девятая) субъединица обозначается символом δ и расположена на внешней стороне F1.
Минорная
– одиночная субъеденица.
Мембранная часть АТФ-синтетазы, представляет собой водо-отталкивающий белковый комплекс, пронизывающий мембрану насквозь и имеющий внутри себя два полуканала для прохождения протонов водорода. Всего в состав комплекса F0 входит одна белковая субъединица типа а
, две копии субъединицы b
, а также от 9 до 12 копий мелкой субъединицы c
. Субъединица а
(молекулярная масса 20 кДа) полностью погружена в мембрану, где образует шесть пересекающих её α-спиральных участков. Субъединица b
(молекулярная масса 30 кДа) содержит лишь один сравнительно короткий погружённый в мембрану α-спиральный участок, а остальная её часть заметно выступает из мембраны в сторону F1 и закрепляется за расположенную на её поверхности субъединицу δ. Каждая из 9-12 копий субъединицы c
(молекулярная масса 6-11 кДа) представляет собой сравнительно небольшой белок из двух водо-отталкивающих α-спиралей, соединённых друг с другом короткой водо-притягивающей петлёй, ориентированной в сторону F1, а все вместе образуют единый ансамбль, имеющий форму погружённого в мембрану цилиндра. Выступающая из комплекса F1 в сторону F0 субъединица γ как раз и погружена внутрь этого цилиндра и достаточно прочно зацеплена за него.
Таким образом, в молекуле АТФазы можно выделить две группы белковых субъединиц, которые могут быть уподоблены двум деталям мотора: ротору и статору.
«Статор»
неподвижен относительно мембраны и включает в себя шарообразный гексамер (αβ)3, находящуюся на его поверхности и субъединицу δ, а также субъединицы a
и b
мембранного комплекса F0.
Подвижный относительно этой конструкции «ротор»
состоит из субъединиц γ и ε, которые, заметно выступая из комплекса (αβ)3, соединяются с погружённым в мембрану кольцом из субъединиц c
.
Способность синтезировать АТФ - свойство единого комплекса F0F1, объедененного с переносом протонов водорода через F0 к F1, в последнем из которых как раз и расположены центры реакции, осуществляющие преобразование АДФ и фосфата в молекулу АТФ. Движущей же силой для работы АТФ-синтетазы является протонный (положительно заряженный) потенциал, создаваемый на внутренней мембране митохондрий в результате работы цепи электронного (отрицательно заряженного) транспорта.
Сила, приводящая в движение «ротор» АТФ-синтетазы, возникает при достижении разности потенциалов между наружной и внутренней сторонами мембраны > 220 10−3 Вольт и обеспечивается потоком протонов, протекающих через специальный канал в F0, расположенный на границе между субъединицами a
и c
. При этом путь переноса протонов включает в себя следующие структурные элементы:
1) Два расположенных на разных осях «полуканала», первый из которых обеспечивает поступление протонов из межмембранного пространства к существенно важным функциональным группам F0, а другой обеспечивает их выход в матрикс митохондрии;
2) Кольцо из субъединиц c
, каждая из которых в своей центральной части содержит протонируемую карбоксильную группу (COOH), способную присоединять H+ из межмембранного пространства и отдавать их через соответствующие протонные каналы. В результате периодических смещений субъединиц с
, обусловленных потоком протонов через протонный канал происходит поворот субъединицы γ, погружённой в кольцо из субъединиц с
.
Таким образом, объединяющая активность АТФ-синтетазы непосредственно связана с вращением её «ротора», при котором поворот субъединицы γ вызывает одновременное изменение конформации всех трёх объединяющих субъединиц β, что в конечном счёте и обеспечивает работу фермента. При этом в случае образования АТФ «ротор» крутится по часовой стрелке со скоростью четыре оборота в секунду, а само подобное вращение происходит точными скачками по 120°, каждый из которых сопровождается образованием одной молекулы АТФ.
Работа АТФ-синтетазы связана с механическими движениями её отдельных частей, что позволило отнести этот процесс к особому типу явлений, названных «вращательным катализом». Подобно тому, как электрический ток в обмотке электродвигателя приводит в движение ротор относительно статора, направленный перенос протонов через АТФ-синтетазу вызывает вращение отдельных субъединиц фактора сопряжения F1 относительно других субъединиц ферментного комплекса, в результате чего это уникальное энергообразующее устройство совершает химическую работу - синтезирует молекулы АТФ. В дальнейшем АТФ поступает в цитоплазму клетки, где расходуется на самые разнообразные энергозависимые процессы. Подобный перенос осуществляется специальным встроенным в мембрану митохондрий ферментом АТФ/АДФ-транслоказой.
АДФ-транслоказа
– пронизывающий внутреннюю мембрану белок, который обменивает вновь синтезированную АТФ на цитоплазматическую АДФ, что гарантирует сохранность фонда внутри митохондрий.
Митохондрии и наследственность.
ДНК митохондрий наследуются почти исключительно по материнской линии. Каждая митохондрия имеет несколько участков нуклеотидов в ДНК, идентичных во всех митохондриях (то есть в клетке много копий митохондриальных ДНК), что очень важно для митохондрий, неспособных восстанавливать ДНК от повреждений (наблюдается высокая частота мутаций). Мутации в митохондриальной ДНК являются причиной целого ряда наследственных заболеваний человека.
3д модель
Дисковери
С англ озвучкой
Немного о дыхании клетки и митохондрии на зарубежном языке
Структура строения
Митохондрии — это микроскопические мембранные органоиды, которые обеспечивают клетку энергией. Поэтому их называют энергетическими станциями (аккумулятором) клеток.
Митохондрии отсутствуют в клетках простейших организмов, бактерий, энтамеб, которые живут без использования кислорода. Некоторые зеленые водоросли, трипаносомы содержат одну большую митохондрию, а клетки сердечной мышцы, мозга имеют от 100 до 1000 данных органелл.
Особенности строения
Митохондрии относятся к двухмембранным органеллам, имеют внешнюю и внутреннюю оболочки, межмембранное пространство между ними и матрикс.
Внешняя мембрана
. Она гладкая, не имеет складок, отграничивает внутреннее содержимое от цитоплазмы. Ширина ее равна 7нм, в составе находятся липиды и белки. Важную роль выполняет порин - белок, образующий каналы во внешней мембране. Они обеспечивают ионный и молекулярный обмен.
Межмембранное пространство
. Величина межмембранного пространства около 20нм. Вещество, заполняющее его по составу сходно с цитоплазмой, за исключением крупных молекул, которые могут сюда проникнуть только путем активного транспорта.

Внутренняя мембрана
. Построена в основном из белка, только треть отводится на липидные вещества. Большое количество белков являются транспортными, так как внутренняя мембрана лишена свободно проходимых пор. Она формирует много выростов – крист, которые выглядят, как приплюснутые гребни. Окисление органических соединений до CO 2 в митохондриях происходит на мембранах крист. Этот процесс кислородзависимый и осуществляется под действием АТФ-синтетазы. Высвобожденная энергия сохраняется в виде молекул АТФ и используется по мере необходимости.
Матрикс
– внутренняя среда митохондрий, имеет зернистую однородную структуру. В электронном микроскопе можно увидеть гранулы и нити в клубках, которые свободно лежат между кристами. В матриксе находится полуавтономная система синтеза белка – здесь расположены ДНК, все виды РНК, рибосомы. Но все же большая часть белков поставляется с ядра, поэтому митохондрии называют полуавтономными органеллами.
Расположение в клетке и деление
Хондриом
– это группа митохондрий, которые сосредоточены в одной клетке. Они по-разному располагаются в цитоплазме, что зависит от специализации клеток. Размещение в цитоплазме также зависит от окружающих ее органелл и включений. В клетках растений они занимают периферию, так как к оболочке митохондрии отодвигаются центральной вакуолью. В клетках почечного эпителия мембрана образует выпячивания, между которыми находятся митохондрии.
В стволовых клетках, где энергия используется равномерно всеми органоидами, митохондрии размещены хаотично. В специализированных клетках они, в основном, сосредоточены в местах наибольшего потребления энергии. К примеру, в поперечно-полосатой мускулатуре они расположены возле миофибрилл. В сперматозоидах они спирально охватывают ось жгутика, так как для приведения его в движение и перемещения сперматозоида нужно много энергии. Простейшие, которые передвигаются при помощи ресничек, также содержат большое количество митохондрий у их основания.
Деление
. Митохондрии способны к самостоятельному размножению, имея собственный геном. Органеллы делятся с помощью перетяжки или перегородок. Формирование новых митохондрий в разных клетках отличается периодичностью, например, в печеночной ткани они сменяются каждые 10 дней.
Функции в клетке
- Основная функция митохондрий – образование молекул АТФ.
- Депонирование ионов Кальция.
- Участие в обмене воды.
- Синтез предшественников стероидных гормонов.
Молекулярная биология – это наука, изучающая роль митохондрий в метаболизме. В них также идет превращение пирувата в ацетил-коэнзим А, бета-окисление жирных кислот.
| Таблица: строение и функции митохондрий (кратко)
|
|---|
| Структурные элементы
| Строение
| Функции
|
| Наружная мембрана
| Гладкая оболочка, построена из липидов и белков
| Отграничивает внутреннее содержимое от цитоплазмы
|
| Межмембранное пространство
| Находятся ионы водорода, белки, микромолекулы
| Создает протонный градиент
|
| Внутренняя мембрана
| Образует выпячивания – кристы, содержит белковые транспортные системы
| Перенос макромолекул, поддержание протонного градиента
|
| Матрикс
| Место расположения ферментов цикла Кребса, ДНК, РНК, рибосом
| Аэробное окисление с высвобождением энергии, превращение пирувата в ацетил-коэнзим А.
|
| Рибосомы
| Объединённые две субъединицы
| Синтез белка
|
Сходство митохондрий и хлоропластов
Общие свойства для митохондрий и хлоропластов обусловлены, прежде всего, наличием двойной мембраны.
Признаки сходства также заключаются в способности самостоятельно синтезировать белок. Эти органеллы имеют свое ДНК, РНК, рибосомы.
И митохондрии и хлоропласты могут делиться с помощью перетяжки.
Объединяет их также возможность продуцировать энергию, митохондрии более специализированы в этой функции, но хлоропласты во время фотосинтезирующих процессов тоже образуют молекулы АТФ. Так, растительные клетки имеют меньше митохондрий, чем животные, потому что частично функции за них выполняют хлоропласты.
Опишем кратко сходства и различия:
- Являются двомембранными органеллами;
- внутренняя мембрана образует выпячивания: для митохондрий характерны кристы, для хлоропластов – тиллакоиды;
- обладают собственным геномом;
- способны синтезировать белки и энергию.
Различаются данные органоиды своими функциями: митохондрии предназначены для синтеза энергии, здесь осуществляется клеточное дыхание, хлоропласты нужны растительным клеткам для фотосинтеза.
Бытует крепко укрепившееся мнение, что выносливость человека связано с тренировкой сердечной мышцы, и что для этого нужно длительное время выполнять невысокую по интенсивности работу.
На самом деле всё не так: выносливость неразрывно связано с митохондриями внутри мышечных волокон. Поэтому тренировка выносливости есть не что иное, как развитие максимального количества митохондрии внутри каждого мышечного волокна.
А т.к. максимальное количество митохондрий ограничено пространством внутри мышечного волокна, то и развитие выносливости ограничено тем количеством мышц, которые присутствуют у конкретного человека.
Короче: чем больше у человека митохондрий внутри конкретных мышечных групп, тем более выносливыми являются эти конкретные мышечные группы.
И самое важное: не существует общей выносливости. Есть только локальная выносливость конкретных мышечных групп.
Митохондрии. Что это такое
Митохондрии – это особенные органеллы (структуры) внутри клеток человеческого организма, которые отвечают за производство энергии для мышечных сокращений. Иногда их называют энергетическими станциями клетки.
При этом процесс производства энергии внутри митохондрий происходит в присутствии кислорода. Кислород делает процесс получения энергии внутри митохондрий максимально эффективным, если сравнивать процесс получения энергии без кислорода.
Топливом для производства энергии могут являются совершенно различные вещества: жир, гликоген, глюкоза, лактат, ионы водорода.
Митохондрии и выносливость. Как это происходит
При мышечном сокращении всегда появляется остаточный продукт. Обычно это молочная кислота – химическое соединение из лактата и ионов водорода.
По мере накопления внутри мышечного волокна (мышечной клетки) ионы водорода начинают вмешиваться в процесс получения энергии для сокращения мышечного волокна. А как только уровень концентрации ионов водорода достигает критической отметки, мышечное сокращение прекращается. И данный момент может свидетельствовать об максимальном уровне выносливости конкретной мышечной группы.
Митохондрии обладают способностью поглощать ионы водорода и перерабатывать их внутри себя.
Получается следующая ситуация. Если внутри мышечных волокон присутствует большое количество митохондрий, то они способны утилизировать и большее количество ионов водорода. А это означает более длительную работу конкретной мышцы без необходимости прекратить усилие.
В идеале, если митохондрий внутри работающих мышечных волокон достаточно для утилизации всего количества образующихся ионов водорода, то такое мышечное волокно становится практически неутомимым и способным продолжать работу до тех пор, пока будет достаточное количество питательных веществ для сокращения мышц.
Пример.
Почти каждый из нас способен длительное время идти быстрым темпом, но довольно скоро бывает вынужден прекратить бег быстрым темпом. Почему так выходит?
При быстрой ходьбе работают т.н. окислительные и промежуточные мышечные волокна. Окислительные мышечные волокна характеризуются максимально возможным количеством митохондрий, грубо говоря, митохондрий там 100 %.
В промежуточных мышечных волокнах митохондрий ощутимо меньше, пусть это будет 50 % от максимального количества. В итоге, постепенно внутри промежуточных мышечных волокон начинают накапливаться ионы водорода, которые должны бы привести к прекращению сокращения мышечных волокон.
Но этого не происходит по причине того, что ионы водорода проникают внутрь окислительных мышечных волокон, где митохондрии без труда справляются с их утилизацией.
В итоге, мы способны продолжать движения до тех пор, пока в организме достаточно гликогена, а также запасов жира внутри работающих окислительных мышечных волокон. Затем мы будем вынуждены сделать отдых для пополнения запасов энергии.
В случае с быстрым бегом в работу, помимо упомянутых окислительных и промежуточных мышечных волокон, включаются и т.н. гликолитические мышечные волокна, в которых почти отсутствуют митохондрии. Поэтому гликолитические мышечные волокна способны работать лишь короткое время, зато крайне интенсивно. Именно таким образом повышается скорость бега.
Потом общее количество ионов водорода становится таким, что всё количество имеющихся там же митохондрий уже не способно утилизировать их. Наступает отказ от выполнения работы предложенной интенсивности.
Но что было бы, если бы все мышечные группы имели внутри себя только окислительные мышечные волокна?
В этом случае мышечная группа с окислительными волокнами становится неутомимой. Ее выносливость становится равной бесконечности (при условии достаточного количества питательных веществ – жиров и гликогена).
Делаем следующий вывод: Для тренировки выносливости первоочередное значение имеет развитие митохондрий внутри рабочих мышечных волокон. Именно благодаря митохондриям достигается выносливость мышечных групп.
Не существует общей выносливости организма, потому что выносливость (способность выполнять работу предложенной интенсивности) связана с присутствием в работающих мышцах митохондрий. Чем митохондрий там больше, тем большую выносливость способны показать мышцы.
Строение и функции ядра растительной клетки.
Ядро
– обязательная часть эукариотической клетки. Это место хранения и воспроизведения наследственной информации. Ядро также служит центром управления обменом веществ и почти всех процессов, происходящих в клетке. Чаще всего в клетках имеется лишь одно ядро, редко - два или несколько. Форма его чаще всего шаровидная или эллипсоидальная. В молодых, особенно меристематических, клетках оно занимает центральное положение, но позднее обычно смещается к оболочке, оттесняемое растущей вакуолью. Снаружи ядро покрыто двойной мембраной – ядерной оболочкой, пронизанной порами (поры ядра - динамичные образования, они могут открываться и закрываться; таким путем может осуществляться регуляция обмена между ядром и цитоплазмой) на краях которых наружная мембрана переходит во внутреннюю. Наружная ядерная мембрана соединяется с мембранными каналами ЭПС. На ней располагаются рибосомы. Внутренняя мембрана может давать впячивания.
Внутреннее содержимое ядра – кариоплазма с погруженными в нее хроматином и ядрышками, и рибосомами. Кариоплазма (нуклеоплазма) представляет собой желеобразный раствор, который заполняет пространство между структурами ядра (хроматином и ядрышками). Она содержит ионы, нуклеотиды, ферменты.
Хроматин – это деспирализованная форма существования хромосом. В деспирализованном состоянии хроматин находится в ядре неделящейся клетке. Хроматин и хромосомы взаимно переходят друг в друга. По химической организации как хроматин, так и хромосомы не отличаются. Химическую основу составляет дезоксирибонуклеопротеин – комплекс ДНК с белками. С помощью белков происходит многоуровневая упаковка молекул ДНК, при этом хроматин приобретает компактную форму.
Ядрышко, обычно шаровидной формы (одно или несколько), не окружено мембраной, содержит фибриллярные белковые нити и РНК. Ядрышки – не постоянные образования, они исчезают в начале деления клетки и восстанавливаются после его окончания. Ядрышки имеются только в неделящихся клетках. В ядрышках происходит формирование рибосом, синтез ядерных белков. Сами же ядрышки образуются на участках вторичных перетяжек хромосом (ядрышковых организаторах).
Ядро – обязательная часть эукариотической клетки. Диаметр ядра колеблется от 5 до 20 мкм. Главная функция ядра – хранение генетического материала в форме ДНК и передача ее дочерним клеткам при клеточном делении. Кроме того, ядро управляет белковыми синтезами, контролирует все процессы жизнедеятельности клетки. (в растительной клетке ядро описал Р.Броун в 1831г., в животной – Т.Шванн в 1838г.).
Химический состав ядра представлен, главным образом, нуклеиновыми кислотами и белками.
Строение и функции митохондрий.
Митохондрии или хондриосомы - «силовые» станции клетки, в них локализована большая часть реакций дыхания (аэробная фаза). В митохондриях происходит аккумуляция энергии дыхания в аденозинтрифосфате (АТФ). Энергия, запасаемая в АТФ, служит основным источником для физиологической деятельности клетки. Митохондрии обычно имеют удлиненную палочковидную форму длиной 4-7 мкм и диаметром 0,5-2 мкм. Число митохондрий в клетке может быть различным от 500 до 1000 и зависит от роли данного органа в процессах энергетического обмена.
Химический состав митохондрий несколько колеблется. В основном это белково-липидные органеллы. Содержание белка в них составляет 60-65%, причем структурные и ферментативные белки содержатся примерно в равной пропорции, а также около 30% липидов. Очень важно, что митохондрии содержат нуклеиновые кислоты: РНК - 1 % и ДНК -0,5%. В митохондриях имеется не только ДНК, но и вся система синтеза белка, в том числе рибосомы.
Митохондрии окружены двойной мембраной. Толщина мембран составляет 6-10 нм. Мембраны митохондрий на 70% состоят из белка. Фосфолипиды мембран представлены фосфатидтилхолином, фосфатидилэтаноламином, а также специфическими фосфолипидами, например, кардиолипином. Мембраны митохондрий не пропускают Н+ и служат барьером для их транспорта.
Между мембранами находится заполненное жидкостью перимитохондриальное пространство. Внутреннее пространство митохондрий заполняет матрикс в виде студнеобразной полужидкой массы. В матриксе сосредоточены ферменты цикла Кребса. Внутренняя мембрана дает выросты - кристы в виде пластин и трубочек, они разделяют внутреннее пространство митохондрий на отдельные отсеки. Во внутренней мембране локализована дыхательная цепь (цепь переноса электронов).
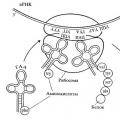 Центральная догма молекулярной биологии Схема центральной догмы молекулярной биологии
Центральная догма молекулярной биологии Схема центральной догмы молекулярной биологии Нил Шубин Вселенная внутри нас: что общего у камней, планет и людей
Нил Шубин Вселенная внутри нас: что общего у камней, планет и людей Майкл Фарадей: биография
Майкл Фарадей: биография